擔杮楍搰傊偺恖椶偺棳擖
丂
妀DNA夝愅偵傛傞撽暥恖偺弌帺
丂擔杮楍搰偵恖椶偑傗偭偰偒偨偺偼偄偮側偺偱偁傠偆偐丅崱夞丄暉搰導偺嶰娧抧奓捤偱敪孈偝傟偨恖崪偐傜妀DNA偑夝撉偝傟丄乽撽暥恖偼傾僕傾偺懠偺抧堟偺恖偨偪偲戝偒偔堎側傞摿挜傪帩偭偰偄偨乿偲傢偐偭偨丅偦傟偽偐傝偐丄尰戙偺擔杮恖偲傕梊憐埲忋偵堘偄偑戝偒偐偭偨偲帵偝傟偨丅偦偺夝撉寢壥傪傕偲偵擔杮楍搰傊偺撽暥恖偺棳擖傪峫偊偰傒傛偆丅
丂2016擭9寧丄崙棫堚揱妛尋媶強側偳偺僌儖乕僾偑乽撽暥恖偺妀僎僲儉傪弶傔偰夝撉偟偨乿偲偡傞榑暥傪愱栧帍偵敪昞偟偨丅偙偺乽撽暥恖乿偲偼丄暉搰導怴抧挰偵偁傞嶰娧抧奓捤偱敪孈偝傟偨俁愮擭慜偺恖崪偱偁傞丅嶰娧抧奓捤偼丄徍榓俀侽擭戙偵侾侽侽懱埲忋偺恖崪偑敪孈偝傟偨撽暥帪戙傪戙昞偡傞奓捤偱偁傞丅尋媶僌儖乕僾偼丄搶嫗戝妛偵曐娗偝傟偰偄偨恖崪丒抝彈俀恖偺墱帟偺撪懁偐傜傢偢偐側俢俶俙傪嵦庢偟丄夝愅偵惉岟偟偨偺偱偁傞丅
丂偦偺暘愅寢壥傪傕偲偵恖椶偺宯摑庽傪昞偡偲師偺傛偆偵側傞丅
| 丂丂妀DNA夝愅偵傛傝敾柧偟偨恖椶偺宯摑庽 丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂 恖椶劒劒劒劒劚劒劒劒劒劒劒劒劒劒劒劒劒劒傾僼儕僇恖 丂丂丂丂丂丂劔 丂丂丂丂丂丂劔丂劕劒劒劒劒劒劒劒劒劒劒劒儓乕儘僢僷恖 丂丂丂丂丂丂劔丂劔 丂丂丂丂丂丂劘劒劜丂劕劒劒劒劒劒劒劒劒劒僷僾傾僯儏乕僊僯傾恖 丂丂丂丂丂丂丂丂劔丂劔 丂丂丂丂丂丂丂丂劘劒劜丂劕劒劒劒劒劒劒劒撽暥恖 丂丂丂丂丂丂丂丂丂丂劔丂劔丂丂丂丂丂丂丂丂伀12%嫟捠DNA偺懚嵼丂 丂丂丂丂丂丂丂丂丂丂劘劒劜丂丂丂丂丂劕劒擔杮恖 丂丂丂丂丂丂丂丂丂丂丂丂劔丂丂丂劕劒劜 丂丂丂丂丂丂丂丂丂丂丂丂劔丂丂丂劔丂劘劒拞崙恖 丂丂丂丂丂丂丂丂丂丂丂丂劔丂劕劒劜丂丂丂 丂丂丂丂丂丂丂丂丂丂丂丂劔丂劔丂劘劒劒劒搶撿傾僕傾恖 丂丂丂丂丂丂丂丂丂丂丂丂劘劒劜 丂丂丂丂丂丂丂丂丂丂丂丂丂丂劘劒劒劒劒劒傾儊儕僇愭廧柉 |
丂夝撉偟偨偺偼嶰娧抧奓捤偺撽暥恖偱偁傞偑丄撽暥帪戙撽暥恖摨巑偺岎棳偑偐側傝妶敪偱偁偭偨偺偱丄DNA偺抧堟奿嵎偼偐側傝彮側偄傕偺偲悇掕偡傞丅尰戙偺擔杮恖偲撽暥恖偑DNA偱墦偔棧傟偰偍傝丄傎偐偺傾僕傾恖偲傕戝偒偔棧傟偰偄傞丅傑偨丄DNA偺撍慠曄堎偵傛傞曄壔偼擭悢偲偲傕偵戝偒偔側傞偺偱丄撽暥恖偑擔杮楍搰偵棳擖偟偰偐傜戝棨偺恖椶偲挿偔妘愨偝傟偰偄偨偙偲傪堄枴偟偰偄傞丅
Y愼怓懱偺堚揱巕夝愅偵傛傞尋媶寢壥
丂僴僾儘僌儖乕僾偲偼丄暘巕恖椶妛偱梡偄傜傟傞丄恖椶偺Y愼怓懱僴僾儘僌儖乕僾乮扨攞孮乯偺暘椶偱偁傞丅暘壔偟偨弴偵A,B,C,D丒丒丒偲T傑偱暘椶偝傟偰偍傝丅擔杮恖偺傒偲偝傟偰偄傞D1b偼僴僾儘僌儖乕僾D偺僒僽僋儗乕僪乮嵶暘婒乯偺堦偮偱丄3枩擭傎偳慜偵擔杮楍搰偱抋惗偟偨偲峫偊傜傟偰偄傞丅尰嵼擔杮恖偺3妱側偄偟4妱偑偙偺僌儖乕僾偵懏偟偰偄傞丅偙偺暘壔偺宯摑傪捛媮偡傞偙偲偵傛偭偰丄撽暥恖偺擔杮楍搰傊偺堏摦宱楬偑柧傜偐偵側傝偮偮偁傞丅
僴僽儘僌儖乕僾傪傕偲偵偟偨恖椶偺堏摦夁掱偺悇掕
丂尰惗恖椶愨柵偺婋婡
丂偙偺乽恑壔偺宯摑庽乿傪傕偲偵偟偰撽暥恖偺棳擖夁掱傪悇掕偡傞偲偵偡傞偲師偺傛偆偵側傞丅乽儂儌丒僒僺僄儞僗乿偲屇偽傟傞尰惗恖椶偼俀侽枩擭慜偵傾僼儕僇偱抋惗偟丄懠偺戝棨偵恑弌偟偨丅14枩擭慜偖傜偄偵傾僼儕僇戝棨傪弌偨偲巚傢傟傞丅拞崙偱敪尒偝傟偨尨惗恖椶偺壔愇(桍峕恖)偺惗懚帪婜傕彮側偔偲傕13.6枩擭慜傑偱偝偐偺傏傝丄僕儍儚搰偱敪尒偝傟偨12.5枩擭慜崰偺帟傕尰惗恖椶偺傕偺偲悇掕偝傟偰偄傞丅偙偺偙偲偐傜丄14枩擭慜偖傜偄偺戞1夞栚偺弌傾僼儕僇偱丄尨惗恖椶偼偐側傝峀斖埻偵搉偭偰奼嶶偟偰偄偨丅
丂偲偙傠偑丄栺11.5枩擭慜偐傜偼嵟屻偺昘壨帪戙丄栺9枩擭慜偐傜偼僂儖儉昘婜偺嵟廔昘婜偑巒傑偭偨丅俈枩擭慜偵偼嵟廔昘婜偺垷昘婜偑巒傑傝丄偙偺帪戙埲崀偼姦椻偱姡憞偟偨婥岓偵側偭偨偺偱偁傞丅
丂奀柺偼尰嵼傛傝傕 100乣130m 傕掅壓偟丄僀僊儕僗偼戝棨偲棨懕偒偲側傝丄僪僀僣杒晹傑偱昘尨偑撿壓丅怷椦抧堟偼僀儀儕傾敿搰傗抧拞奀増娸偺傢偢偐側抧堟偩偗偲側偭偨丅搶撿傾僕傾廃曈傕棨抧壔偟丄僗儞僟儔儞僪偑抋惗偟丄擔杮楍搰傕儐乕儔僔傾戝棨偲抧懕偒偲側傞丅偦偺忋丄7枩5000乣4000擭慜偵僗儅僩儔搰偺杒晹偵偁傞僩僶壩嶳偺戝暚壩偑偁偭偨丅偙偺暚壩偵傛傞壩嶳偺搤偱悽奅偺婥壏偼1乣5亷掅壓偟丄偦傟偑1000擭掱傕懕偄偨偲偝傟偰偄傞丅丅暚壩捈屻偺2乣3擭偼15亷傕掅壓偟杒敿媴偱昘愥抧懷奼戝偟偨丅傾僼儕僇偐傜僀儞僪傪挻偊偰搶撿傾僕傾偵恑弌偟偨尰惗恖椶偼丄偙偺僩僶壩嶳偺暚壩偺塭嬁偱戝検巰柵偟偨傛偆偱偁傞丅
丂偙偺壩嶳暚壩偵傛傝丄傾僼儕僇戝棨偼彊乆偵庈擬壔偟丄戝澺榀偲側傝丄僒僴儔偺憪尨偼晄栄偺嵒敊偲壔偟丄傾僼儕僇偼撿杒偵暘抐偝傟偨丅僸僩偺堚揱巕偺懡條惈偼戝宆椶恖墡偵斾傋嬃偔傎偳掅偄丅偙傟偼丄僩僶壩嶳偺戝暚壩偵傛傝丄傾僼儕僇偵巆偭偨恖乆傕丄僒僴儔偑嵒敊壔偟偨偨傔摦暔偼傕偪傠傫丄怉暔偝偊摼傜傟側偔側傝丄嶨怘壔偱惗偒墑傃偨偑丄2000恖傎偳偺愨柵悺慜忬懺偵娮偭偨丅偙傟偑尨場偱堚揱揑側懡條惈偺懡偔偑幐傢傟偨偨傔偲峫偊傜傟偰偄傞丅
丂弌傾僼儕僇
丂偙偺忬懺偵娮偭偨恖椶偵僴僾儘僌儖乕僾CT傪掕媊偯偗傞曄堎偑7枩擭慜偛傠僄僠僆僺傾偐傜僗乕僟儞偺曈傝偵偄偨堦恖偺抝惈偵惗偠偨丅偙偺Y愼怓懱忋偺DNA墫婎攝楍偺曄堎傪丄堦晹偺傾僼儕僇恖埲奜偺慡恖椶偺抝惈偼庴偗宲偄偱偄傞丅僴僾儘僌儖乕僾乽A宯摑乿偲乽B宯摑乿偼偦傟埲慜偵丄暘婒偟偰偍傝丄傾僼儕僇偐傜弌偰偄側偄丅偦偺屻丄65000擭慜偛傠僴僽儘僌儖乕僾CT偺巕懛偱偁傞搶傾僼儕僇偺僩僁儖僇僫屛偺搶杒晬嬤偵廧傫偱偄偨堦恖偺抝惈偵僴僽儘僌儖乕僾DE傊偺曄堎偑婲偙偭偨丅傑偨丄傎傏摨偠偙傠CF宯摑傊偺曄堎傕婲偙偭偨丅偙偺DE宯摑偼D1丄D2丄E偺宯摑偵暘壔偟丄D1,E,CF宯摑偺恖乆偼摨偠偙傠丄婥岓偺曄摦偵廬偄丄傾僼儕僇戝棨偺搶晹丄尰嵼偺僕僽僠偐傜僄儕僩儕傾偺曈傝偐傜峠奀乮僶僽丒僄儖丒儅儞僨僽奀嫭乯傪搉偭偰丄尰嵼偺僀僄儊儞撿晹偵摓払偟丄弌傾僼儕僇傪壥偨偟偨丅偙偺巕懛偨偪偼僒僴儔嵒敊埲撿埲奜偺慡悽奅偵揱攄偟斏怋偟偰偄偭偨偲巚傢傟傞丅
丂僴僾儘僌儖乕僾C偺恖乆偺堏摦
丂僴僽儘僌儖乕僾C偺恖乆偼60000擭慜偛傠惣撿傾僕傾偺尰僀儔儞晅嬤偱抋惗偟偨丅C偼娫傕側偔C1偲側傝丄C1偼C1a偲C1b偵暘婒偟偨丅 C1a偺堦晹偑儓乕儘僢僷曽柺偵堏摦偟丄C1a偺堦晹偲C1b偑僀儞僪偐傜僗儞僟儔儞僪乮搶撿傾僕傾偵偁偭偨戝棨乯偵堏摦偟偨丅C1a偼拞墰傾僕傾偺曽偵恑弌偟50000擭慜偛傠C2偵曄堎偟偨丅
丂C2偼儅儞儌僗傪捛偭偰僔儀儕傾偵廧傒拝偄偨丅儅儞儌僗偼嫄戝惗暔偱偁傝丄堦摢巇棷傔傞偲敿擭偖傜偄偺娫怘椏偵崲傜側偄傎偳偺傕偺偱偁偭偨丅抁偄壞偵儅儞儌僗傪巇棷傔丄搤偼偦傟傪怘傋偰惗偒偰偄偔偲偄偆宍偲側偭偰偄偨丅嫄戝惗暔儅儞儌僗偼丄僌儖乕僾偱嫤椡偟側偄偲巇棷傔傞偙偲偑偱偒側偄丅僌儖乕僾偱徖抧偵捛偄崬傫偱丄嵶愇恘傪梡偄偰巇棷傔偨傕偺偲巚傢傟傞丅嵶愇恘偼崪偺椉懁偵崟梛愇偺攋曅傪庢傝晅偗偨傕偺偱丄儅儞儌僗偺崪傪娧偔傎偳偺埿椡偑偁偭偨丅
丂偙偺摉帪偺戙昞揑堚愓偼僶僀僇儖屛斎偺儅儕僞堚愓偱偁傞丅23000擭偛傠傑偱塰偊偰偄偨C2a偺僌儖乕僾偱偁傞丅23000擭慜偛傠嵟廔昘婜偺嵟姦椻婜偑嬤偯偄偰偄偨丅抁偄壞傕側偔側傝丄惗妶偑嬯偟偔側傝丄僔儀儕傾偐傜撿傗搶偵堏摦偟偰偄偭偨丅搶偵堏摦偟偨恖乆偺堦晹偼C2a1偺僌儖乕僾偲側傝丄20000擭慜偛傠搥傝晅偄偨儀乕儕儞僌奀傪搉偭偰丄杒傾儊儕僇戝棨偵搉偭偨丅偝傜偵堦晹偼姃懢偵堏摦偟丄姃懢偐傜杒奀摴偵堏傝廧傒C2a2偺僌儖乕僾偲側偭偨丅嵟姦椻婜偺偙傠丄搥寢偟偨捗寉奀嫭傪搉傝杮廈偵擖傝崬傓偙偲偲側偭偨丅偙偺宯摑偼擔杮屌桳偺宯摑偱偁傞丅撿偵堏摦偟偨恖乆偺堦晹偼C2b偲側傝丄拞崙戝棨傗挬慛敿搰偵掕廧偟偰偄偭偨丅
丂C1a偺恖乆偼C1b偺恖乆偲偲傕偵僗儞僟儔儞僪偵堏傝廧傫偱偄偨丅昘壨帪戙偲偼尵偊抔偐偔丄怘椏偵偼嬯楯偟側偐偭偨偱偁傠偆丅偟偐偟丄嵟廔昘婜偑廔傢傝丄奀悈柺偑忋徃偟丄師戞偵僗儞僟儔儞僪偑悈杤偟偰偒偨丅恖乆偼慏偵忔偭偰堏摦偡傞偙偲偲側偭偨丅C1a偺恖乆偼崟挭偵忔偭偰杒忋偟丄壂撽偵偨偳傝拝偄偨丅17000擭傎偳慜偺偙偲偱偁傞丅峘愳恖偱偁傞丅
丂偙偺崰偼崟挭偑嬨廈偺撿懁傪棳傟偰偍傝丄摉帪偺慏偱偼嬨廈偵偨偳傝拝偔偙偲偑偱偒側偐偭偨丅偟偐偟丄15000擭傎偳慜偵懳攏奀嫭偑峀偑傝丄崟挭偐傜懳攏奀棳偑暘棳偡傞傛偆偵側傝丄廙偱撿嬨廈偵棳傟拝偔偙偲偑偱偒傞傛偆偵側傝丄撿嬨廈偵偨偳傝拝偄偨丅C1a1宯摑偺恖暔偼擔杮楍搰屌桳偱偁傝丄尰戙擔杮恖偵5%傎偳懚嵼偟偰偄傞丅C1a宯摑偼40000擭傎偳慜偵敪尰偟偰偄傞偑丄偦偺巕宯摑偺奼嶶偑巒傑偭偨偺偑12000擭傎偳慜偐傜偱偁傝丄偦偺娫偵挿偄僊儍僢僾偑偁傞丅偙傟傛傝丄擔杮楍搰棳擖帪婜偑晄妋掕偲側傞偑丄奼嶶偑巒傑傞彮偟慜偵棳擖偟偨偲峫偊偰偄傞丅C1a1宯摑偵嬤偄C1a2偑儓乕儘僢僷傗傾儖儊僯傾偵懡偄丅偦偺娫偺抧堟偵懚嵼偣偢丄偦偺宱楬偑摿掕偱偒側偄丅嬤偄宯摑偺C1b偺懡偄僗儞僟儔儞僪偐傜棃偨偲悇掕偟偰偄傞偑丄恖悢傕彮側偔丄僗儞僟儔儞僪偵巆偭偨C1a1宯摑偼愨柵偟偨偺偱偼側偄偩傠偆偐丅
丂峘愳恖偺僴僽儘僌儖乕僾偼妋擣偝傟偰偄側偄偑丄弌搚偟偨恖崪偺崪奿偑僆乕僗僩儔儕傾偺尨廧柉偲帡偰偄傞偺偱丄僗儞僟儔儞僪偐傜棃偨宯摑偲巚傢傟丄C1a1宯摑偲悇掕偡傞丅
丂僴僽儘僌儖乕僾D偺恖乆偺堏摦
丂D2宯摑偼傾僼儕僇偐傜弌偰偍傜偢丄傾僼儕僇偐傜弌偨D1宯摑偼6.5枩擭慜偵DE宯摑偐傜暘偐傟偨偲巚傢傟傞丅D1宯摑偼拞墰傾僕傾偺僠儀僢僩偵堏摦偟丄D1a宯摑偲側偭偨丅53000擭傎偳慜偵D1a2偺宯摑偑僠儀僢僩偐傜撿偵堏摦偟偨偺偑傾儞僟儅儞彅搰偵堏傝丄D1a2b偲側偭偨丅搶偵堏摦偟偨偺偑丄38000擭傎偳慜偵挬慛敿搰偐傜擔杮楍搰偵忋棨偟偨偲巚傢傟傞丅擔杮楍搰忋棨屻D1a2a偺宯摑偲側偭偨丅偙偺宯摑偼擔杮楍搰偺傒偵偟偐懚嵼偣偢丄尰戙擔杮恖偺39%傪愯傔偰偍傝丄峜幒偵偮側偑偭偰偄傞丅D2宯摑偼悽奅揑偵偼悢偑彮側偄宯摑偱丄偦偺椞堟偼偮側偑偭偰偍傜偢丄搶傾僕傾偱偼擔杮楍搰偵偟偐偄側偄丅偙傟偼丄柵朣偟偨傕偺偲峫偊傜傟傞丅
丂DE宯摑偺堚揱巕偼YAP堚揱巕乮恄偺堚揱巕乯偲傕屇偽傟丄嬑曌丄拵偺壒傪鉟楉偩偲巚偆姶惈丄帺暘傪幪偰偰偱傕懠偺恖偺偨傔偵摦偔偙偲偑偱偒傞偲偄偆惈幙偼偙偺堚揱巕偵傛傞傕偺偲尵傢傟偰偄傞丅偦偺偨傔偵丄拞崙戝棨傗挬慛敿搰偱偼懠柉懓偲偺峈憟偵偍偄偰憟傢偢丄柵傏偝傟偰偟傑偭偨偺偱偼側偄偐偲峫偊傜傟傞丅
丂恖椶偺擔杮楍搰傊偺棳擖夁掱
丂D1a2偺宯摑偺恖乆偼懳攏奀嫭傪墇偊偨偲巚傢傟傞偑丄38000擭慜偵偳偺傛偆偵偟偰懳攏奀嫭傪墇偊偨偺偱偁傠偆偐丅
丂
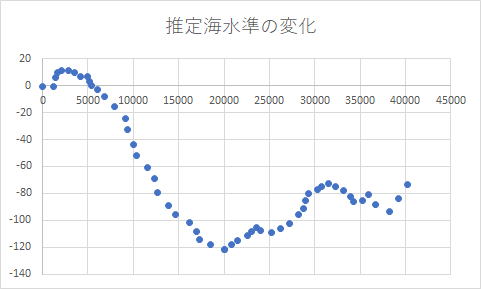
丂忋偺僌儔僼偼40000擭慜埲崀偺奀悈柺偺曄埵偺悇掕抣傪昞偟偨丅墶幉偑崱偐傜壗擭慜偐傪帵偟丄廲幉偼尰嵼偺奀悈柺傪0m偲偟偨偲偒偺偦偺摉帪偺悇掕奀悈柺傪帵偟偰偄傞丅奀悈柺偑壓偑偭偰偄傞帪偼姦椻婜偵偁偨傞偲偒偱偁傞丅懳攏奀嫭偺嵟怺晹偼悈怺210m偱丄悈怺200m埲怺偺晹暘偼奀姌偲側偭偰偍傝丄暆5km挿偝60km傎偳偱偁傞丅奀姌偼挭棳偵傛偭偰怹怘偝傟偨晹暘偺偙偲偱偁傞丅嵟廔昘婜嵟姦椻婜偺20000擭慜偵嵟傕奀柺偑壓偑偭偰偄傞偑丄偙偺帪偱傕懳攏奀嫭偼懚嵼偟偰偄傞丅偙偺帪偼暆15km傎偳偱偁傠偆丅偟偐偟丄怹怘偝傟偨偲偄偆偙偲傪峫偊傟偽丄怹怘偡傞慜偼偮側偑偭偰偄偨壜擻惈偑峫偊傜傟傞丅30000擭慜偐傜15000擭慜傑偱偼奀悈柺偑崱傛傝100m埲忋壓偑偭偰偄傞偺偱寖偟偔怹怘偟偰偄傞偙偲偑峫偊傜傟傞偑丄偦偺慜偺38000擭傎偳慜偵傕100m傎偳奀悈柺偑壓偑偭偰偄傞帪婜偑偁傞丅偙偺帪偼傑偩怤怘偟偰偄側偄偲傕峫偊傜傟丄挬慛敿搰偲懳攏偑棨懕偒偱偁偭偨壜擻惈傕峫偊傜傟傞丅D1a2a偺恖乆偑擔杮楍搰偵棳擖偟偨帪婜偲廳側傞偺偱丄偍偦傜偔丄偙偺帪婜偵棨懕偒偱偁偭偨偺偱偼側偄偩傠偆偐丅
丂僴僽儘僌儖乕僾O宯摑偺棳擖
丂偙偺宯摑偺恖乆偼撽暥帪戙斢婜偐傜栱惗帪戙偵偐偗偰棳擖偟偨恖乆偺宯摑偱偁傞丅僗儞僟儔儞僪曽柺偵岦偐偭偨C1宯摑偺恖乆偐傜僀儞僪偱暘壔偟偨偲悇掕偝傟偰偄傞丅C1宯摑偺恖乆偲偲傕偵僗儞僟儔儞僪偵堏摦偟丄4枩擭傎偳慜偵僗儞僟儔儞僪偐傜拞崙戝棨偵堏摦偟偨恖乆偐傜O宯摑偺暘壔偑婲偙偭偨偲峫偊傜傟偰偄傞丅擔杮楍搰偱偼O1a丄O1b丄O2a丄O2b偑懡偔丅慡懱偺53%傎偳傪愯傔傞丅
丂O1a偼峕撿恖偺慯宯偱丄擔杮偺3亾傎偳傪愯傔傞丅戜榩尨廧柉偑66%偱嵟傕懡偔丄搶撿傾僕傾偵20%傎偳丄尰戙拞崙偵10%傎偳懚嵼偟偰偄傞丅慯娍愴憟偱慯偼婭尦慜202擭偵娍偵攕傟偨丅偙偺帪偵摝朣偟偰偒偨恖乆偲巚傢傟傞丅撿嬨廈偱捈愙朄偺惢揝傪偟偰偄偨偲悇掕偝傟傞埳淨檉懜偑偙偺宯摑偱偼側偄偐偲巚傢傟傞丅
丂O1b偼O1b1偲O1b2偵暘偐傟傞丅O1b1偼峕撿偺墇宯偱丄擔杮偺1亾傎偳傪愯傔傞丅墇偼婭尦慜334擭偵慯偵攕傟丄偦偺恖乆偼儀僩僫儉丄儅儗乕僔傾丄僶儕搰丄儃儖僱僆搰丄僕儍儚搰側偳偵摝朣偟丄擔杮偵傕嬐偐偵傗偭偰棃偨丅墇宯偼拞崙偵傕15亾偺斾棪偱懚嵼偡傞丅
丂O1b2偼屶宯偱丄擔杮慡懱偺33亾傎偳傪愯傔傞丅婭尦慜473擭偵屶偼墇偵攕傟丄杒曽乮彊廈乯偵摝朣偟偨丅挬慛敿搰撿晹傗拞崙杒搶晹偵傕摝朣丒掕廧偟偨丅屻偺懢攲偺巕懛偺宯摑乮埳淨戻懜丒嬬搝崙乯傗丄挬慛敿搰偐傜傗偭偰偒偨恖乆偑偙傟偵奩摉偡傞偲巚傢傟傞丅慺岬歫懜傗弌塤墹挬傕偙偺宯摑偱偼側偄偩傠偆偐丅
丂O2a偼墿壨宯偱丄擔杮偺20亾傎偳傪愯傔傞丅拞崙偵55亾丄挬慛敿搰偵44亾丄儅儗乕僔傾丄僞僀丄僠儀僢僩丄儀僩僫儉側偳搶撿傾僕傾偵傕懡偄丅恅偺巒峜掗偵捛傢傟擔杮楍搰偵傗偭偰偒偨恖乆偵偙偺宯摑偑懡偐偭偨偲巚傢傟丄栱惗帪戙偺杒嬨廈彅崙乮搝崙丒埳搒崙乯偺恖乆偑偙傟偵奩摉偡傞偲巚傢傟傞丅
丂O2b偼擔杮偵1.5%傎偳懚嵼偟丄挿峕拞丒壓棳堟偱敪惗偟偨偲峫偊傜傟傞丅挿峕壨岥堟弌恎偺恅彊暉偑偙偺宯摑偱偼側偄偩傠偆偐丅
丂儈僩僐儞僪儕傾僴僾儘僌儖乕僾偵偮偄偰
丂Y愼怓懱僴僾儘僌儖乕僾偼抝宯偱宲彸偝傟傞丅偙傟偵懳偟偰儈僩僐儞僪儕傾僴僾儘僌儖乕僾偼彈宯偱宲彸偝傟傞丅儈僩僐儞僪儕傾僴僾儘僌儖乕僾傪傑偲傔偰傒傛偆丅乮乽DeNA偑DNA夝愅両擔杮恖偺慶愭偺儖乕僣傪偨偳傞乿傛傝乯
| 僌儖乕僾 | 斾棪 | 婲尮 | 暘晍 | 旛峫 |
| D | 35 | 拞崙拞晹 | 拞崙丒挬慛敿搰 | 堫嶌偲偲傕偵栱惗帪戙偵棳擖 |
| B | 13 | 拞崙撿晹 | 懢暯梞丒撿暷 | 撿曽偐傜棳擖偟偨撽暥恖 |
| M7 | 13 | 僗儞僟儔儞僪 | 擔杮撈摿 | 撿曽偐傜棳擖偟偨撽暥恖 |
| G | 7 | 拞崙杒晹 | 僔儀儕傾 | 杒曽偐傜棳擖偟偨撽暥恖丒栱惗恖 |
| A | 7 | 僔儀儕傾 | 搶傾僕傾丒傾儊儕僇戝棨 | 杒曽傛傝棳擖偟偨撽暥恖 |
| N9 | 7 | 儐乕儔僔傾杒晹 | 僇儉僠儍僢僇丒搶杒傾僕傾 | 撽暥恖丒栱惗恖 |
| F | 5 | 搶撿傾僕傾 | 搶撿傾僕傾 | 挬慛敿搰傛傝棳擖 |
| Z | 1 | 拞崙杒晹 | 僔儀儕傾丒杒墷 | |
| M8a | 1 | 拞崙杒晹 | 拞崙娍柉懓 | 栱惗恖 |
| C | 1 | 拞崙杒晹 | 拞墰傾僕傾丒傾儊儕僇戝棨 | 栱惗帪戙偵棳擖 |
丂儈僩僐儞僪儕傾僴僾儘僌儖乕僾偼曄壔偑寖偟偄偺偱丄Y愼怓懱傎偳柧妋偵堏摦宱楬偑暘偐傜側偄丅撽暥帪戙偵傕栱惗帪戙偵傕棳擖偟偨摨偠僌儖乕僾傕偁傞偲巚傢傟傞丅
丂
| 丂僩僢僾傊丂丂栚師傊丂 | |
| 師 | 擔杮楍搰偺恖乆 |
| 娭楢 | 閌懍擔懜旘懰崙朘栤 枴彌崅旻崻柦抋惗 弌塤墹挬抋惗 |